
 HOME
HOME Cell-free tech
Cell-free tech Research
Research Paper outline
Paper outline Member
Member Publication
Publication Funds
Funds Contact us
Contact us Link
Link Photo
Photo 講義用資料
講義用資料
AGIA tag system: 高親和性抗DRD1ウサギモノクローナル抗体を用いた新規アフィニティタグ
翻訳後修飾されるアミノ酸を含んでいない細胞内で発現したタンパク質の高感度検出・免疫沈降に最適化している
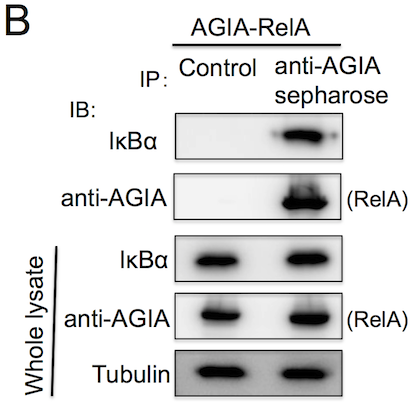 FLAGタグやMycタグに代表される特異的ペプチドと抗体からなるアフィニティタグシステムは生化学や細胞生物学の実験において必須のツールとして使われています。これまでに主に5種類のタグが利用されてきましたが、細胞内で翻訳後修飾されるアミノ酸を含む、細胞内の他のタンパク質と反応するなど、いくつかの改善すべき点がありました。2015年にコムギ無細胞技術を用いて得られた抗ドーパミン受容体DRD1抗体
FLAGタグやMycタグに代表される特異的ペプチドと抗体からなるアフィニティタグシステムは生化学や細胞生物学の実験において必須のツールとして使われています。これまでに主に5種類のタグが利用されてきましたが、細胞内で翻訳後修飾されるアミノ酸を含む、細胞内の他のタンパク質と反応するなど、いくつかの改善すべき点がありました。2015年にコムギ無細胞技術を用いて得られた抗ドーパミン受容体DRD1抗体 Takeda, et al.,は、10-10 M程度の高親和性を有するウサギモノクローナル抗体であったことから、この抗体を用いた高機能な新規タグシステム(AGIA tag system)の開発を進めました。その結果、n Mの高親和性を示し、これまでのタグでは検出が難しかったタンパク質の検出に成功しました。また、細胞内の発現タンパク質の免疫染色や免疫沈降が高感度に行え、またAlphaScreen技術への応用も可能であることが分かりました。さらにタグ配列の1つのアミノ酸を置換したところ、ペプチドを用いたタンパク質の溶出精製に成功しました。以上のことから、AGIA tag systemは、これまでに使われているタグシステムを超える高感度かつ高い特異性を示す新規アフィニティタグシステムであることが分かりました。
Takeda, et al.,は、10-10 M程度の高親和性を有するウサギモノクローナル抗体であったことから、この抗体を用いた高機能な新規タグシステム(AGIA tag system)の開発を進めました。その結果、n Mの高親和性を示し、これまでのタグでは検出が難しかったタンパク質の検出に成功しました。また、細胞内の発現タンパク質の免疫染色や免疫沈降が高感度に行え、またAlphaScreen技術への応用も可能であることが分かりました。さらにタグ配列の1つのアミノ酸を置換したところ、ペプチドを用いたタンパク質の溶出精製に成功しました。以上のことから、AGIA tag systemは、これまでに使われているタグシステムを超える高感度かつ高い特異性を示す新規アフィニティタグシステムであることが分かりました。
 Paper
Paper![]()
標的タンパク質の責任ユビキチンE3リガーゼをハイスループットに同定する技術
コムギ無細胞系を基盤としたプロテインアレイは責任ユビキチンE3リガーゼの同定に適している
 タンパク質のユビキチン化は真核生物に保存された翻訳後修飾であり、標的となるタンパク質の寿命や機能、局在などを決定する細胞内の重要な因子です。タンパク質がユビキチン化される際には、ユビキチンE3リガーゼ(E3)が標的となるタンパク質に特異的に結合してユビキチン化することが分かっています。また、20,000種類以上とも言われるヒトのタンパク質の少なくとも1/3は細胞内でユビキチン化されると言われており、これだけのタンパク質に対応するために、E3もヒトでは600種類以上存在しています。この複雑さゆえ、どのタンパク質をどのE3がユビキチン化するかの組み合せは、その多くが未解明なままでした。
タンパク質のユビキチン化は真核生物に保存された翻訳後修飾であり、標的となるタンパク質の寿命や機能、局在などを決定する細胞内の重要な因子です。タンパク質がユビキチン化される際には、ユビキチンE3リガーゼ(E3)が標的となるタンパク質に特異的に結合してユビキチン化することが分かっています。また、20,000種類以上とも言われるヒトのタンパク質の少なくとも1/3は細胞内でユビキチン化されると言われており、これだけのタンパク質に対応するために、E3もヒトでは600種類以上存在しています。この複雑さゆえ、どのタンパク質をどのE3がユビキチン化するかの組み合せは、その多くが未解明なままでした。
我々は250種類のヒトおよびマウスからなるE3のタンパク質アレイを作製し、これらのE3と標的タンパク質の結合を、384穴プレート上で高感度かつハイスループットに探索するアッセイ技術を開発しました。また、がん抑制遺伝子であるp53をモデルとしたプレスクリーニングにより、p53に結合するE3をいくつも見出しました。この中には、既にp53をユビキチン化することが報告されているMDM2などの他、これまで報告されていなかったE3も含まれており、その中でもRNF6やDZIP3は実際にp53をユビキチン化することが分かりました。これらのことから、このアッセイ系はこれまで未知であったE3と標的タンパク質の組み合せを見出す新たな技術となることが期待されます。
 Paper
Paper![]()
創薬ターゲットGPCRに対するモノクローナル抗体の新しい作製技術を確立
コムギ無細胞系は創薬ターゲットのGPCRタンパク質の合成や抗GPCR抗体の作成・選抜に適している
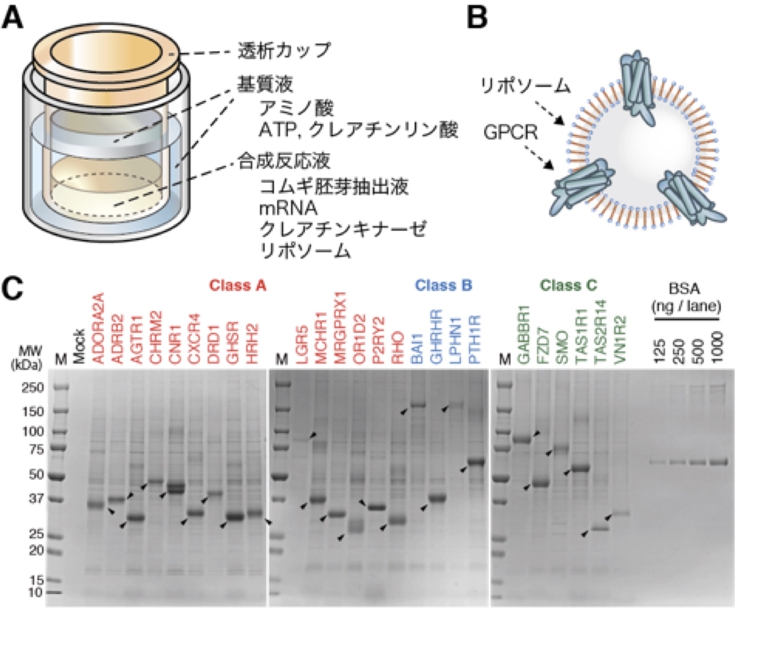 愛媛大学発の技術であるコムギ無細胞タンパク質合成技術を応用し,GPCR(Gタンパク質共役受容体)タンパク質の大量合成法と抗GPCR抗体選抜法を開発し,新たな抗GPCRモノクローナル抗体作製技術を確立しました。
愛媛大学発の技術であるコムギ無細胞タンパク質合成技術を応用し,GPCR(Gタンパク質共役受容体)タンパク質の大量合成法と抗GPCR抗体選抜法を開発し,新たな抗GPCRモノクローナル抗体作製技術を確立しました。
GPCRは最も重要な創薬ターゲットのひとつと知られ,その機能解析や活性制御のためにはGPCRを特異的に認識する抗体が必要でした。しかし,従来の発現系では抗体作製に必要な大量のGPCRの調製が困難であり,また有用な抗体を効率的に選抜する手法がなかったため,GPCRに対する抗体開発は難しいのが現状でした。本技術を用いれば有用な抗GPCR抗体の作製が容易になり,GPCRを対象とした基礎研究のみならずGPCRを標的とした抗体医薬開発への貢献が期待できます。
 詳細資料
詳細資料 Paper
Paper![]()
植物のチロシンキナーゼを世界で初めて発見
植物特有のCDPK-related kinase (CRK)ファミリーは、植物細胞中のタンパク質のチロシン残基をリン酸化している。
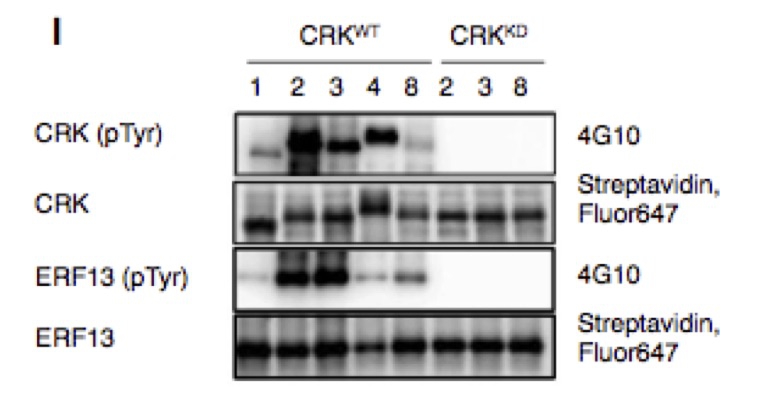 細胞内の情報伝達を制御している機構として、タンパク質のリン酸化があります。タンパク質のリン酸化を触媒する酵素は大きく分けて2種類あり、セリン/スレオニン残基をリン酸化するセリン/スレオニンキナーゼ、チロシン残基をリン酸化するチロシンキナーゼです。動物細胞の増殖や分化などにおいてチロシンキナーゼが重要な役割を担っており、現在、市販されている多くの抗がん剤はチロシンキナーゼを標的としています。しかし、植物には動物で見つかったチロシンキナーゼと似た遺伝子がなかったため、植物にはチロシンキナーゼは無いと長い間いわれてきました。しかし、最近のプロテオミクス解析などで、植物細胞内からチロシン残基がリン酸化されたタンパク質が沢山見つかり、植物にもきっとチロシンキナーゼがあるに違いないと考えられてきました。
細胞内の情報伝達を制御している機構として、タンパク質のリン酸化があります。タンパク質のリン酸化を触媒する酵素は大きく分けて2種類あり、セリン/スレオニン残基をリン酸化するセリン/スレオニンキナーゼ、チロシン残基をリン酸化するチロシンキナーゼです。動物細胞の増殖や分化などにおいてチロシンキナーゼが重要な役割を担っており、現在、市販されている多くの抗がん剤はチロシンキナーゼを標的としています。しかし、植物には動物で見つかったチロシンキナーゼと似た遺伝子がなかったため、植物にはチロシンキナーゼは無いと長い間いわれてきました。しかし、最近のプロテオミクス解析などで、植物細胞内からチロシン残基がリン酸化されたタンパク質が沢山見つかり、植物にもきっとチロシンキナーゼがあるに違いないと考えられてきました。
そこで我々は、シロイヌナズナ植物のプロテインキナーゼを759種類合成し、自分自身のチロシン残基をリン酸する酵素を38種類見出し、その中から、別のタンパク質分子のチロシン残基をリン酸化する酵素、CDPK-related kinase (CRK)を発見しました。CRKファミリーに属するタンパク質は、細胞内で転写因子ERF13やチューブリンのチロシン残基をリン酸化し、CRKの遺伝子が変異している植物個体では、チューブリンのリン酸化チロシンは劇的に減っていました。これらの成果は、世界初の植物チロシンキナーゼの発見といえます。現在、植物におけるリン酸化チロシンの生物学的意味の解明に挑戦しています。
 Paper
Paper![]()
新しいミオシンIIの不活性化機構の発見
ミオシンホスファターゼはアポトーシス時の不可逆的にされる
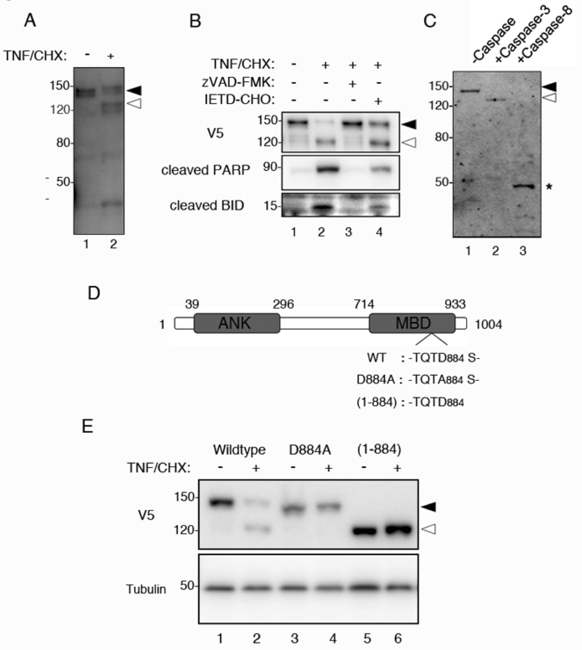 ミオシンIIは動物細胞の移動や分裂、形態変化等の多様な細胞運動に関与する重要なモーター蛋白質です。通常、細胞内のミオシンIIはRhoキナーゼによるリン酸化とミオシンホスファターゼによる脱リン酸化を恒常的に受けており、その活性は時期特異的に調節されています。しかし、細胞がアポトーシスを起こす際にはRhoキナーゼの著しい活性化がおこり、ミオシンIIが過剰に活性化されてしまうため、アポトーシスに特異的な激しい細胞膜ブレッビングが誘導されることが広く知られています。その一方で、ミオシンIIの不活性化の役割を担っているミオシンホスファターゼがアポトーシス時にどのように働いているのかは、これまで明らかにされていませんでした。
ミオシンIIは動物細胞の移動や分裂、形態変化等の多様な細胞運動に関与する重要なモーター蛋白質です。通常、細胞内のミオシンIIはRhoキナーゼによるリン酸化とミオシンホスファターゼによる脱リン酸化を恒常的に受けており、その活性は時期特異的に調節されています。しかし、細胞がアポトーシスを起こす際にはRhoキナーゼの著しい活性化がおこり、ミオシンIIが過剰に活性化されてしまうため、アポトーシスに特異的な激しい細胞膜ブレッビングが誘導されることが広く知られています。その一方で、ミオシンIIの不活性化の役割を担っているミオシンホスファターゼがアポトーシス時にどのように働いているのかは、これまで明らかにされていませんでした。
そこで我々は、ヘテロ三量体であるミオシンホスファターゼ分子の中で、ミオシンII結合能を持つMYPT1に着目して、アポトーシス時における機能解析を行いました。その結果、MYPT1はアポトーシス時にリン酸化とカスパーゼ3による切断を受けることによってミオシンII結合能を失い、細胞内でミオシンIIと共局在できなくなることがわかりました。それに伴いミオシンホスファターゼのミオシンII脱リン酸化能が低下することを見出しました。また、MYPT1をsiRNAでノックダウンするとアポトーシス誘導時における死細胞の割合が増加することも明らかになりました。
 Paper
Paper![]()
TRB3はプロカスパーゼ3を核に局在させ、アポトーシスを回避させる
小胞体ストレスにおける新しいアポトーシス回避機構モデル
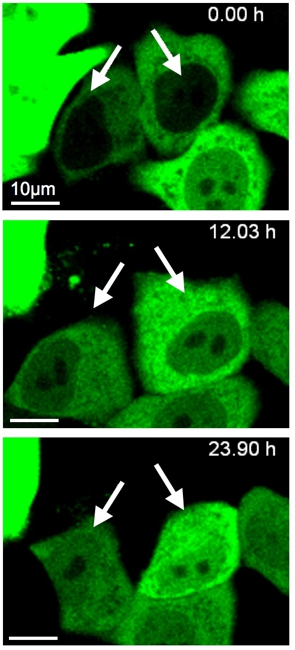 細胞は様々なストレスを受けています。特に小胞体(ER)ストレスは、膜タンパク質や分泌タンパク質が正しくフォールディングできない場合や、小胞体内での糖鎖付加が阻害される等、頻繁に起こるストレスといわれています。一過的な小胞体ストレスの場合、細胞はタンパク質合成抑制などストレス応答を行いストレスを解消とします。しかし、容認できる以上のストレスを受けた場合、細胞はアポトーシスにより“死”を迎えます。この小胞体ストレス応答において、一時的にアポトーシスを回避する機構は未解明な部分が多いのが現状です。本研究では、小胞体ストレスで誘導されるTRB3というタンパク質が、細胞質に局在しているアポトーシスに必須なカスパーゼ3前駆体(procaspase-3)を核内へ誘導し、その結果、カスパーゼ3活性化を抑制し、アポトーシスを回避していることを見出しました。
細胞は様々なストレスを受けています。特に小胞体(ER)ストレスは、膜タンパク質や分泌タンパク質が正しくフォールディングできない場合や、小胞体内での糖鎖付加が阻害される等、頻繁に起こるストレスといわれています。一過的な小胞体ストレスの場合、細胞はタンパク質合成抑制などストレス応答を行いストレスを解消とします。しかし、容認できる以上のストレスを受けた場合、細胞はアポトーシスにより“死”を迎えます。この小胞体ストレス応答において、一時的にアポトーシスを回避する機構は未解明な部分が多いのが現状です。本研究では、小胞体ストレスで誘導されるTRB3というタンパク質が、細胞質に局在しているアポトーシスに必須なカスパーゼ3前駆体(procaspase-3)を核内へ誘導し、その結果、カスパーゼ3活性化を抑制し、アポトーシスを回避していることを見出しました。
 Paper
Paper
細胞質局在カスパーゼ3前駆体(proCASP3-EGFP)は、アポトーシス誘導を促進するが、核局在カスパーゼ3前駆体(proCASP3-NLS-EGFP)は、アポトーシス刺激に影響しない。赤く染まる細胞は、コントロールの核局在RFP。
![]()
植物がもつタンパク質フォスファターゼの網羅的生化学解析
シロイヌナズナはリン酸化チロシンを脱リン酸化する酵素を合計4種類もっている
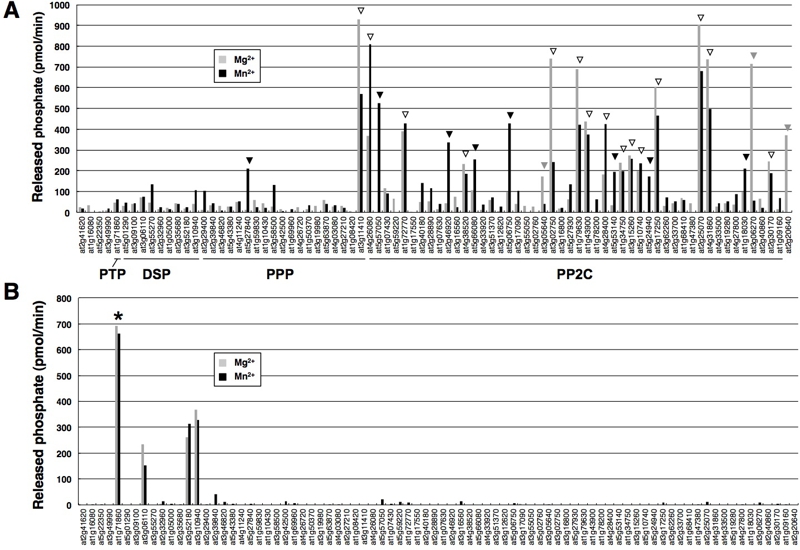 植物は、タンパク質のリン酸化残基を認識しリン酸基を外す脱リン酸化活性を有する酵素(タンパク質フォスファターゼ)を100種類近く持っている。しかし、それらの生化学的解析はほとんど進んでいない。そこで私達は、理化学研究所が整備したシロイヌナズナ完全長cDNAライブラリーの中から、82種類のタンパク質フォスファターゼ遺伝子を選び出し、コムギ無細胞タンパク質合成技術でタンパク質を合成し、生化学的解析を行いました。その結果、30種類のタンパク質フォスファターゼが単独で活性をもつ事がわかりました。その内、21種類のタンパク質フォスファターゼの活性測定は世界初でした。種々のタンパク質フォスファターゼの活性化には、マグネシウムイオンもしくはマンガンイオンのどちらを必要とする、基質として、リン酸化セリン/スレオニンもしくはリン酸化チロシンを脱リン酸化するか、などが明らかとなり、今後、植物のタンパク質フォスファターゼの機能解析のための基本となるデータを得ることが出来ました。
植物は、タンパク質のリン酸化残基を認識しリン酸基を外す脱リン酸化活性を有する酵素(タンパク質フォスファターゼ)を100種類近く持っている。しかし、それらの生化学的解析はほとんど進んでいない。そこで私達は、理化学研究所が整備したシロイヌナズナ完全長cDNAライブラリーの中から、82種類のタンパク質フォスファターゼ遺伝子を選び出し、コムギ無細胞タンパク質合成技術でタンパク質を合成し、生化学的解析を行いました。その結果、30種類のタンパク質フォスファターゼが単独で活性をもつ事がわかりました。その内、21種類のタンパク質フォスファターゼの活性測定は世界初でした。種々のタンパク質フォスファターゼの活性化には、マグネシウムイオンもしくはマンガンイオンのどちらを必要とする、基質として、リン酸化セリン/スレオニンもしくはリン酸化チロシンを脱リン酸化するか、などが明らかとなり、今後、植物のタンパク質フォスファターゼの機能解析のための基本となるデータを得ることが出来ました。
 Paper
Paper![]()
カスパーゼ8はIL21シグナルを負に制御する
カスパーゼ8によるインターロイキン21(IL21)シグナルの負の制御機構モデル
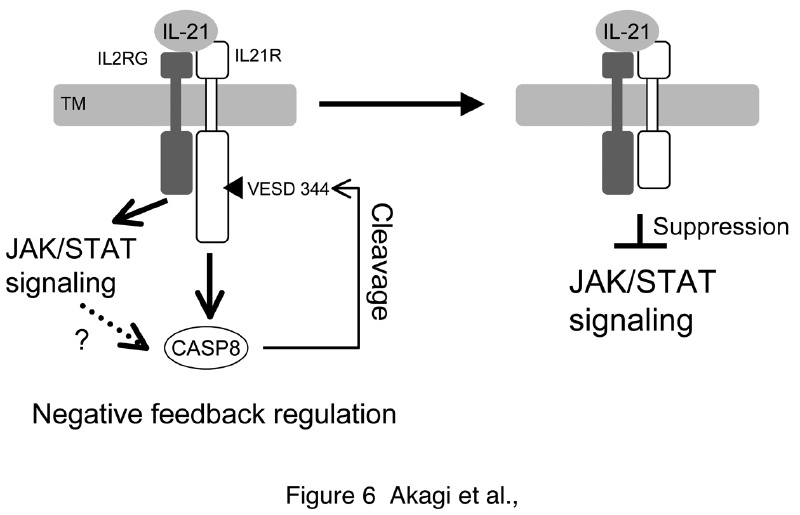 カスパーゼ8前駆体(procaspase-8)は、細胞膜内側近くに存在し、TNFαやFasといったアポトーシス誘導シグナルにより活性化し、カスパーゼ3などの活性化を誘導し、細胞死を引き起こします。しかし、近年、細胞死以外にも、マクロファージ分化など、血球系の分化などに関わっている可能性が指摘されています。そこで、我々は407種類のシングル膜貫通領域をもつ膜タンパク質(sTMP)からなるNCtaggedタンパク質ライブラリーを作成し、カスパーゼ8の基質探索を行いました。その結果、3つのsTMPがカスパーゼ8の基質であること、中でも、IL21受容体がカスパーゼ8のみならず、カスパーゼ3にも切断されることを見出しました。STAT3リン酸化を指標にIL21シグナルを調べたところ、カスパーゼ8/3切断IL21受容体は、STAT3のリン酸化を誘導できないことから、カスパーゼ8は、IL21受容体の切断を介して、IL21シグナルを負に制御することがわかりました。
カスパーゼ8前駆体(procaspase-8)は、細胞膜内側近くに存在し、TNFαやFasといったアポトーシス誘導シグナルにより活性化し、カスパーゼ3などの活性化を誘導し、細胞死を引き起こします。しかし、近年、細胞死以外にも、マクロファージ分化など、血球系の分化などに関わっている可能性が指摘されています。そこで、我々は407種類のシングル膜貫通領域をもつ膜タンパク質(sTMP)からなるNCtaggedタンパク質ライブラリーを作成し、カスパーゼ8の基質探索を行いました。その結果、3つのsTMPがカスパーゼ8の基質であること、中でも、IL21受容体がカスパーゼ8のみならず、カスパーゼ3にも切断されることを見出しました。STAT3リン酸化を指標にIL21シグナルを調べたところ、カスパーゼ8/3切断IL21受容体は、STAT3のリン酸化を誘導できないことから、カスパーゼ8は、IL21受容体の切断を介して、IL21シグナルを負に制御することがわかりました。
 Paper
Paper![]()
難問!! 膜タンパク質の合成・精製への挑戦
N末端に蛍光タンパク質(GFP)を融合した膜タンパク質の密度勾配遠心分離の様子
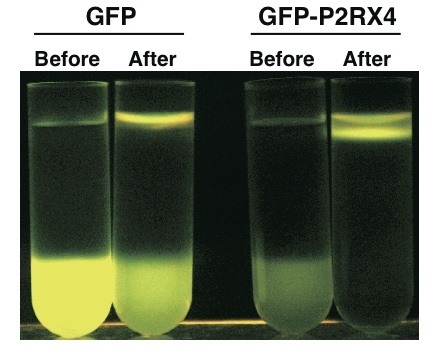 様々な環境変化やホルモンなどは、基本的に細胞膜上の受容体と感知もしくは結合し、細胞内の情報伝達系を介して、遺伝子の発現や酵素活性を変化させます。そのため、受容体がどの様に細胞内へ情報伝達を行っているのかを解明することは非常に重要な研究です。しかし、膜タンパク質の合成は非常に難しく、これまでの技術では、機能をもった膜タンパク質を、人工的に合成する技術はありませんでした。我々の無細胞センターの戸澤教授の研究成果により、コムギ無細胞タンパク質合成系に人工膜であるリポソームを添加することにより、植物の膜タンパク質の1種、リン酸トランスポーターが活性を保持したまま合成することに成功しました(もっと詳しく)。そこで、我々は、他の膜タンパク質の合成にもこの技術が応用できるか、特に、薬開発に必要な膜タンパク質の代表例であるGPCRタンパク質の合成に挑戦しました。また、膜タンパク質は合成だけでなく、水に溶けないため、膜タンパク質精製は、ほとんど不可能なくらい困難な作業です。我々は、リポソームに取り込まれた膜タンパク質を、リポソームごと精製することに成功し、これで、今まで困難であった膜タンパク質の合成・精製するための技術ができました。40種類以上の膜タンパク質がこの技術で合成できたため、非常に汎用性が高い可能性があります。
様々な環境変化やホルモンなどは、基本的に細胞膜上の受容体と感知もしくは結合し、細胞内の情報伝達系を介して、遺伝子の発現や酵素活性を変化させます。そのため、受容体がどの様に細胞内へ情報伝達を行っているのかを解明することは非常に重要な研究です。しかし、膜タンパク質の合成は非常に難しく、これまでの技術では、機能をもった膜タンパク質を、人工的に合成する技術はありませんでした。我々の無細胞センターの戸澤教授の研究成果により、コムギ無細胞タンパク質合成系に人工膜であるリポソームを添加することにより、植物の膜タンパク質の1種、リン酸トランスポーターが活性を保持したまま合成することに成功しました(もっと詳しく)。そこで、我々は、他の膜タンパク質の合成にもこの技術が応用できるか、特に、薬開発に必要な膜タンパク質の代表例であるGPCRタンパク質の合成に挑戦しました。また、膜タンパク質は合成だけでなく、水に溶けないため、膜タンパク質精製は、ほとんど不可能なくらい困難な作業です。我々は、リポソームに取り込まれた膜タンパク質を、リポソームごと精製することに成功し、これで、今まで困難であった膜タンパク質の合成・精製するための技術ができました。40種類以上の膜タンパク質がこの技術で合成できたため、非常に汎用性が高い可能性があります。
 Paper
Paper![]()
植物がもつ自己リン酸化活性型キナーゼの探索
N末端をビオチン化した植物キナーゼの自己リン酸化を抗リン酸化Ser/Thr抗体で検出する技術の原理
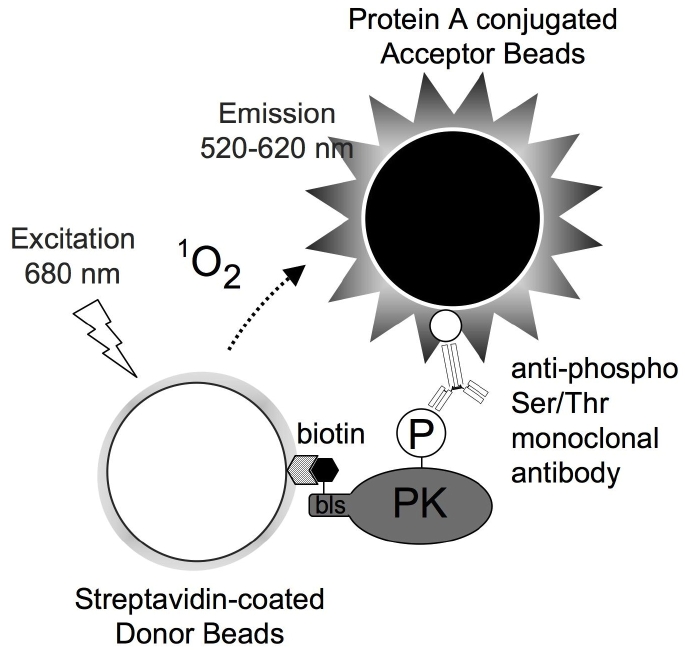 生物は、細胞内に、外的要因や内的要因の変化を的確に伝える情報伝達系を持っている。環境変化やホルモンなどは、基本的に細胞膜上の受容体と感知もしくは結合し、細胞内の情報伝達系を介して、遺伝子の発現や酵素活性を変化させます。そのため、細胞内情報伝達は、細胞増殖や細胞死、細胞分裂など、ほぼ全ての生命現象を制御しています。 プロテインカイネース(通称キナーゼ)は、タンパク質をリン酸化し、構造変化やタンパク質の複合体形成などを誘導するため、細胞内情報伝達の主要経路の1つと考えられています。動物では、解析が進んでおり、抗がん剤ターゲットの多くは、キナーゼです。しかし、植物には、そのゲノム上に動物の2倍ほど(千種類以上)の種類のキナーゼが見つかっているにも関わらず、植物は非常に解析が遅れているのが現状です。遺伝子配列は似ているけれども、本当にこの遺伝子はキナーゼ活性をもっているのかわからない?という様な状況です。遺伝子がキナーゼ活性をもっているかどうか調べるには、遺伝子を鋳型にタンパク質を合成し、タンパク質の活性を調べる必要があります。しかし、これまでの技術では、何百種類もある沢山のキナーゼタンパク質を合成するのが大変でしたので、解析がなかなか進みませんでした。ここで、私達が開発してきたコムギ無細胞タンパク質合成技術の出番です。我々は理化学研究所植物科学センターの篠崎教授がもつ植物(シロイヌナズナ)完全長cDNAクローンの中から、数百種類のキナーゼタンパク質を合成することに成功しました。そこで、キナーゼ活性を有するかどうかの1つの指標として、多くのキナーゼがもつ自己リン酸化活性を調べました。その結果、759種類のシロイヌナズナキナーゼタンパク質の中から、179種類のキナーゼが自己リン酸化活性を有していること、つまりキナーゼ活性を有しているがわかりました。植物にいくつか自己リン酸化活性で自分自身を活性しているため、これは活性制御機構を考える上で指標となります。
生物は、細胞内に、外的要因や内的要因の変化を的確に伝える情報伝達系を持っている。環境変化やホルモンなどは、基本的に細胞膜上の受容体と感知もしくは結合し、細胞内の情報伝達系を介して、遺伝子の発現や酵素活性を変化させます。そのため、細胞内情報伝達は、細胞増殖や細胞死、細胞分裂など、ほぼ全ての生命現象を制御しています。 プロテインカイネース(通称キナーゼ)は、タンパク質をリン酸化し、構造変化やタンパク質の複合体形成などを誘導するため、細胞内情報伝達の主要経路の1つと考えられています。動物では、解析が進んでおり、抗がん剤ターゲットの多くは、キナーゼです。しかし、植物には、そのゲノム上に動物の2倍ほど(千種類以上)の種類のキナーゼが見つかっているにも関わらず、植物は非常に解析が遅れているのが現状です。遺伝子配列は似ているけれども、本当にこの遺伝子はキナーゼ活性をもっているのかわからない?という様な状況です。遺伝子がキナーゼ活性をもっているかどうか調べるには、遺伝子を鋳型にタンパク質を合成し、タンパク質の活性を調べる必要があります。しかし、これまでの技術では、何百種類もある沢山のキナーゼタンパク質を合成するのが大変でしたので、解析がなかなか進みませんでした。ここで、私達が開発してきたコムギ無細胞タンパク質合成技術の出番です。我々は理化学研究所植物科学センターの篠崎教授がもつ植物(シロイヌナズナ)完全長cDNAクローンの中から、数百種類のキナーゼタンパク質を合成することに成功しました。そこで、キナーゼ活性を有するかどうかの1つの指標として、多くのキナーゼがもつ自己リン酸化活性を調べました。その結果、759種類のシロイヌナズナキナーゼタンパク質の中から、179種類のキナーゼが自己リン酸化活性を有していること、つまりキナーゼ活性を有しているがわかりました。植物にいくつか自己リン酸化活性で自分自身を活性しているため、これは活性制御機構を考える上で指標となります。
 Paper
Paper